Дослідницькі роботи і проєкти
Визначення таксономічного статусу кримської популяції жабоп’явки алжирської

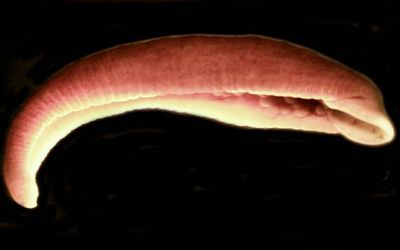
У процесі написання навчального дослідницького проєкта з біології на тему "Визначення таксономічного статусу кримської популяції жабоп’явки алжирської" учень 11 (10) класу ліцею визначає таксономічну належність кримського екземпляру Batracobdella algira та робить його опис. В учнівському проєкті автор розглядає шляхи видоутворення жабоп’явки алжирської та її потенціал розвитку у Криму.
Докладніше про роботу:
В запропонованій науково-дослідницькій роботі з біології у 11 (10 класі) визначається таксономічний статус кримської популяції жабоп’явки алжирської, формуються та представляються результати проведеного дослідження, які дозволяють визначити таксономічний статус кримської популяції Batracobdella algira. В роботі виконується дослідження стану кримської популяції жабоп’явки алжирської, формуються висновки.
Протягом творчої і дослідницької роботи над проєктом про таксономічний статус кримської популяції жабоп’явки алжирської" було проведене дослідження довідкової літератури, за допомогою якої учень 11 класу провів філогенетичний аналіз послідовностей ДНК жабоп’явок з різних ділянок ареалу виду та порівняв кримські й туніські екземпляри жабоп’явок.
В рамках міні-проєкту морфологічні й молекулярні філогенетичні дослідження показали, що кримську й іспанську популяції Batracobdella algira слід вважати окремими, новими для науки видами. Попереднє уявлення про широкий ареал Batracobdella algira, що охоплює Середземномор’я і прісні водойми басейну Чорного моря, не відповідає результатам нашого дослідження та вимагає перегляду.
Зміст
Вступ
1. Видоутворення та його потенціал у Криму.
1.1. Шляхи видоутворення.
1.2. Потенційна цікавість кримського півострова.
1.3. Batracobdella algira.
2. Матеріали і методи.
3. Результати дослідження та їх обговорення.
3.1. Філогенетичне дослідження.
3.2. Морфологічне дослідження.
3.3. Стан кримської популяції.
Висновки
Список використаних джерел
Додатки
Вступ
Питання таксономічної належності виду є дуже важливимм. Тільки розуміючи систематичне положення виду, можна робити правильні висновки щодо нього та його ролі в природних екосистемах, а також правильно організувати його збереження та захист. Крім того, це дозволяє правильно описувати його у джерелах, правильно розуміти ареал його поширення та еволюцію цього виду.
Правильне визначення таксономічного статусу є основним завданням таксономії та систематики, а також важливим розділом зоології. Жабоп’явка алжирська (Batracobdella algira) є видом родини Пласкі п’явки (Glossiphoniidae), який вперше був описаний у Алжирі французьким зоологом Альфредом Мокен-Тандоном, після чого його знаходили в багатьох місцях (Іспанія, Сардинія) включно з Кримом та вивчали цей вид [9].
Така доволі значна поширеність цього виду одразу викликає багато питань щодо таксономічної належності його окремих, часто дуже віддалених популяцій, які ізольовані важко проникними географічними бар’єрами.
Метою роботи є визначення таксономічної належності кримського екземпляру Batracobdella algira та його опис.
Для досягнення поставленої мети нами вирішувались наступні завдання:
- Дослідити питання за літературними джерелами
- Провести філогенетичний аналіз послідовностей ДНК жабоп’явок з різних ділянок ареалу виду.
- Порівняти кримські й туніські екземпляри жабоп’явок.
- Визначити таксономічний статус кримської популяції Batracobdella algira
Об’єкт дослідження: екземпляри Batracobdella algira з Кримського півострова і Північної Африки.
Предмет дослідження: дані філогенетичного аналізу, морфологічний опис.
Для досягнення поставленої мети були використані наступні методи дослідження: біоінформаційні методи (пошук нуклеотидних послідовностей BLAST, вирівнювання нуклеотидних послідовностей, методи побудови філогенетичних дерев Neighbor Joining та Maximum Likelihood), а також методи морфологічних досліджень і мікрофотографії.
Наукова новизна. Морфологічні й молекулярні філогенетичні дослідження показали, що кримську й іспанську популяції Batracobdella algira слід вважати окремими, новими для науки видами. Попереднє уявлення про широкий ареал Batracobdella algira, що охоплює Середземномор’я і прісні водойми басейну Чорного моря, не відповідає результатам нашого дослідження та вимагає перегляду.
Практичне значення отриманих результатів. Таксономічне визначення видів є важливим завданням зоології. Крім того, правильна таксономічна ідентифікація дозволяє точніше визначити природоохоронний статус кримських жабоп’явок і розробити рекомендації та вказівки щодо збереження цього виду та загалом правильно вказувати його у джерелах. До того ж, на основі цих даних можна краще розуміти процесу еволюції для цього виду, повніше розуміти процеси його видоутворення та розповсюдження.
Також Є.І. Лукін у 30 томі серіального видання «Фауна України» каже про те, що цікавим об’єктом дослідження також можуть бути гірудини різних видів п’явок, бо вони можуть по різному впливати на організм, а, значить, використовуватись у різних аспектах гірудотерапії (наприклад, лікувати різними гірудинами різні хвороби) [5]. Тож, правильне розуміння видової належності також може бути корисним і для медицини.
Шляхи видоутворення
Видоутворення – це виникнення нових біологічних видів та зміна старих. Воно спричинене еволюцією та виникає внаслідок сукупної дії різних факторів: мутацій, рекомбінації, природного, добору, дрейфу генів тощо.
У природі існують декілька шляхів видоутворення:
- Дивергентне або справжнє видоутворення, яке, в свою чергу, ділиться на: алопатрічний (ключовий фактор – географічна ізоляція), перипатичний (ключовий фактор – «ефект засновника»), парапатричний (ключовий фактор – досить незначна можливість контактування з первісною популяцією) та симпатричний типи (ключовий фактор – екологічна та / або репродуктивна ізоляція), а також видоутворення шляхом поліплоїдії. Цей шлях характеризується наявністю будь-якої (географічної, екологічної) ізоляції [1].
- Шляхом гібридизації. Цей шлях характеризується відсутністю ізоляції, але наявністю міжвидового схрещування та мутацій, які роблять нащадків від такого розмноження не стерильними [2].
- Філетичне видоутворення, коли весь ареал поступово однаково еволюціонує і всі популяції однаково змінюються під дією часу.
Кожен шлях видоутворення важливий і грав свою роль в утворенні того чи іншого виду. Втім, вважається, що найбільша роль відведена саме алопатрічному типу [3]. На відміну від інших типів, основну роль у алопатрічному типі грає географічна ізоляція. Вона є найвагомішим рушієм видоутворення завдяки різким змінам умов існування внаслідок розділення популяцій, що зрештою зумовлює видоутворення.
Досить сильно може різнитись швидкість видоутворення. Наприклад, деякі бактерії можуть еволюціонувати вже за 8-10 тижнів, втім, зазвичай для цього потрібно куди більше часу, часто мільйони років. На швидкість видоутворення впливає багато факторів, починаючи від ступеня ізольованості й сили тиску відбору, закінчуючи швидкістю закріплення мутацій у геномі.
Ми припускаємо, що у випадку нашого об’єкта дослідження найбільший вплив також мав алопатрічний тип видоутворення. Отже, ми зосередимо свою увагу саме на ньому. Для цього типу важливою умовою є формування географічної ізоляції через дію якої-небудь фактору (переселення, зсув материків, формування островів тощо), внаслідок чого відбувається одразу і репродуктивна ізоляція, і різняться умови, по-різному діє дрейф генів, популяції припиняють обмінюватись генами, різні умови чинять різний тиск природного відбору, і через сукупність всіх цих факторів з плином часу формується спадкова репродуктивна ізоляція, що можна вважати початком існування нового виду.
Потенціальна цікавість Кримського півострова
З точки зору видоутворення, Крим є дуже цікавим місцем досліджень. З материковою частиною України його поєднує лише відносно вузький Перекопський перешийок, що зумовлює певну географічну ізоляцію. Кримський півострів розташований південніше інших областей України, що ще більше посилює різницю в умовах. Навіть порівнявши склад фауни та флори Криму зі складом сусідніх частин України, стає зрозуміло, що видовий склад Криму досить відрізняється і процеси видоутворення тут йшли (і йдуть) особливо активно.
Відособленість Кримського півострова добре видна на прикладі ендемічних видів. Для жодної іншої частини України не властивий такий рівень ендемізму, як Криму. Наприклад, тільки тут живуть такі комахи, як слимакоїд кримський (Carabus (Procerus) scabrosustauricus) та ізофія кримська (Isophyataurica) – її ареал навіть менший за сам півострів.
Вважається, що найбільше ендеміків живе на південному березі, що ще більше підкреслює унікальність фауни Криму). Це підтверджує нещодавнє дослідження мого наукового керівника, що фауна Криму є дослідженою далеко не повністю, а процеси видоутворення там йдуть досить активно серед представників родини Глоткових п’явок [8].
Ендемічні види є й серед рослин. Наприклад, чист кримський (Cistustauricus) та роговик Біберштайна (Cerastiumbiebersteinii). Ми також робимо припущення, що наш об’єкт досліджень є ендеміком Криму та відокремився від початкового виду за вікаріантним типом видоутворення, тобто, перетворення та (або) заміщення початкового виду.
Batracobdella algira
Досить детальний опис жабоп’явки алжирьскої (а точніше, її кримської популяції) дав Є.І. Лукін у 30 томі серіального видання «Фауна України». Опис приводжу нижче: «Морфологічні дані: Розміри. Довжина тіла звичайно не перевищує 17-18 мм; при максимальному розтягненні довжина становить 24 мм, ширина – до 5 мм. В зборах, проведених в Криму в 1958 р., більшість п’явок досягала довжини 7 мм, при ширині 4 мм; найбільш крупні мали в довжину 14 мм, а в ширину 4 мм.
Форма тіла. Тіло видовжене, помірно звужується до переднього кінця, який трохи розширений і тупий. Бокові краї тонко зазублені. Задній присосок круглий, відносно великий, діаметр його майже дорівнює найбільшій ширині тіла (при середньому витягуванні тіла п’явки). Ділянка, що з’єднує присосок з тілом, коротка, сильно звужена.
Поверхня тіла. Спинна сторона трохи випукла і вкрита багатьма дрібними сосочками різної величини. Більші з цих сосочків розміщені в шість повздовжніх паралельних рядів. Слід відзначити, що ці ряди виявлені не досить чітко (не так яскраво, як, наприклад, у Glossiphoniacomplanata). Черевна сторона пласка, рівна.
Забарвлення тіла. Колір спинної сторони зеленувато-коричневий. Чим старша п’явка, тим більше в її забарвленні переважає коричневий пігмент і вона стає зовсім бурою. Оскільки коричневий пігмент більш стійкий, у фіксованих екземплярах у забарвлені переважають коричневі тони. Передня частина тіла п’явок зеленіша, ніж задня. При стисненні п’явки її забарвлення здається більш темним, коричнювато-бурим, при розтягнені, навпаки, краще виявляється зелена пігментація. При більш уважному розгляді можна помітити, що на спинній стороні цієї п’явки проходить 14 коричневих ліній, між якими йдуть більш вузькі світлі, зеленуваті лінії.
У живих п’явок на спині помітні шість повздовжніх рядів жовтих плям, які збігаються з названими вище рядами сосочків. На спинній стороні в місцях статевих отворів є темна пляма. На задньому присоску розташовані також темні плями. Черевна сторона тіла темно-сіра. Тіло малопрозоре або зовсім непрозоре.
Очей двоє, вони великі, неправильної, трикутної або круглястої форми, досить віддалені одне від одного й розташовані на третьому кільці. Очі добре помітні у молодих екземплярів, у дорослих же п’явок в міру нагромадження в шкірі пігментів очі важче побачити. Але після просвітлення п’явок у гліцерині у переважної більшості дорослих особин очі стають помітні.
Шлунок має сім пар відростків, які сильно розширюються до периферії тіла. Передні відростки найменші і гірше помітні, ніж всі інші. Останні відростки найдовші і звисають назад.
Кільчастіть. Склад сомітів такий: І : 1; ІІ : 1; ІІІ : 2; IV-XXIV : 3; XXV : 2 або 1; XXVI : 1; XXVII : 3 або 4.
Статеві отвори розміщені так: чоловічій – між 28- і 29-м кільцями, жіночій – між 30- і 31-м. Між двома отворами – 2-2,5 кільця.
Еколого-фізіологічні дані: Живлення. Описувані п’явки паразитують на різних земноводних, в Криму – на Ranaridibunda. За спостереженнями, проведеними в Криму, батракобдели зустрічаються виключно на дорослих жабах. Вони прикріплюються до них найчастіше в місцях з’єднання ніг з тулубом (з черевної сторони) і на плавальних перетинках; рідше вони трапляються на нижній поверхні голови та в місці колінного з’єднання. У вільному стані п’явки трапляються дуже рідко. Отже, їх можна вважати майже постійними ектопаразитами земноводних. У Криму з однієї жаби знімали до 23 п’явок.
Розмноження. Яйця зовсім круглі (близько 1,5 мм в діаметрі). П’явки з потомством, очевидно, зустрічаються головним чином або виключно у вільному стані. П’явки, які щойно розвинулися з яєць, досягають в довжину 3 мм, забарвлення їх світле, коричнювате, задній присосок відносно тіла більші розміри, ніж у дорослих екземплярів.
Поширення у водоймах. У Криму B. algira живе в різноманітних водоймах, де у великій кількості зустрічаються жаби (канавах, ставках, цементованих басейнах, болотах, стариках річок, у річках).
Географічне поширення: B. algiraзнайдена в Північній Африці, на Піренейському півострові, Балеарських островах, о. Корсиці та в Палестині; в СРСР вона зазначена тільки для Криму, де зустрічається в масовій кількості в околицях Севастополя, Бахчисарая, Сімферополя, Соколиного, Ялти та ін. Певних даних щодо поширення B.
algiraпоки що немає. Цікаво, що досі Batracobdellaalgiraі Batracobdellapaludosaне були відмічені в одній і тій же місцевості і, можливо, що їх ареали взаємно виключають один одного. В Західній Європі межа, що відокремлює два ареали, можливо, збігається з Піренеями, а в Криму вона проходить десь у північній частині півострова. Бажано перевірити (шляхом ретельного огляду дорослих жаб), чи не трапляється B. algiraу водоймах Чорноморського узбережжя за межами Криму» [5].
Матеріали і методи
Дослідження проводилися на екземплярах B. algira, зібраних моїм науковим керівником у 2008 році у Криму біля селища Перевального. Крім того, був досліджений зразок B. algiraз Тунісу, який був зібраний д-ром Раджа Бенахмед Еп Аджилі (Raja Benahmed Ep Ajili) у 2007 році.
П’явок знімали з жаб, які були спіймані у водоймах, після чого фіксували у 96° спирту.
Далі, для проведення молекулярних досліджень, брали зразок ДНК з бокової частини п’явки, після чого моїм науковим керівником виділялась ДНК за наступною методикою: додавались 180 мкл розчину Lysis Solution T, потім 20 мкл розчину протеїнази К до зразка. Далі інгредієнти змішувались за допомогою вортексу, щоб зразок повністю занурився. Потім зразок інкубували за температури 55°С до повного розчинення тканин. Розчинення зазвичай займає 3-6 год. Після завершення розчинення додавали 20 мкл розчину RNase A Solution та інкубували протягом 2 хв за кімнатної температури.
Далі додавали до зразка 200 мкл Lysis Solution C та ретельно розмішували у вортексі (протягом 15 секунд), однорідність суміші є важливою умовою для ефективного лізису. Потім бралась зв’язувальна колона та 2 мл збірна пробірка, додавались 500 мкл Column Preparation Solution до зв’язувальної колонки та центрифугувалось за 12000 g протягом 1 хв. Відфільтрована рідина зливалась, але лишалась зв’язувальна колонка та пробірка.
Додавали 200 мкл етанолу (95-100°) до лізату (однорідної суміші, після додавання Lysis Solution C), ретельно перемішувалось за допомогою вортексу протягом 5-10 секунд, велике значення має гомогенність розчину. Увесь вміст пробірки з лізатом переносилась в попередньо оброблену зв’язувальну колонку з попередніх маніпуляцій, потім проводилось центрифугування за 6500 g протягом 1 хв. Збірна пробірка з осадом відкидалась, а зв’язувальна колонка поміщалась в нову 2-мілілітрову збірну пробірку.
Перед першим використанням концентрат Wash Solution Concentrate розбавлявся етанолом, як описано в інструкції, щодо його приготування. Далі цей розбавлений концентрат у кількості 500 мкл додавався до зв’язувальної колонки та центрифугували протягом 1 хв за 6500 g. Рідина зливалась, а збірна пробірка залишалась. Додавалось ще 500 мкл розчину Wash Solution Concentrate до зв’язувальної колонки, центрифугувалось 3 хв з максимальною швидкістю (12000-16000 g), щоб висушити зв’язувальну колонку. Перед вимиванням ДНК зв’язувальна колонка мала бути вільною від етанолу.
Далі колонка ще раз центрифугували 1 хв за максимальної швидкості, щоб видалити усі залишки етанолу. Збірна пробірка, яка містить рідину, відкидалась і зв’язувальна колонка поміщалась в нову 2-мілілітрову збірну пробірку. Далі додавались 200 мкл Elution Solution безпосередньо в центр зв’язувальної колонки, центрифугували протягом 1 хв за 6500 g, щоб вимити ДНК. Для підвищення ефективності вимивання інкубувалось 5 хв за кімнатної температури після додавання вимивального розчину, далі центрифугували.
Потім шляхом ПЛР виділявся мітохондріальний ген першої субодиниці цитохромоксидази. ПЛР дозволяє отримати багато молекул потрібного саме нам гена, завдяки підбору специфічних праймерів до цього гену та його ампліфікації. Процедура ПЛР була наступною: до пробірки додавались термостійкі полімерази, сам генетичний матеріал, праймери, та йони магнію, після чого пробірка клалась в ампліфікатор.
Програма була наступною: один раз 3 хв розігріву за температури 94 °С, наступні операції циклічно повторювались 30 разів, 45 секунд плавлення (розділення на два ланцюги) ДНК за температури 94 °С, 45 секунд за температури 48 °С, 1 хв за температури в 72°С і останній нагрів за 72 °С, який повторюється 1 раз впродовж 1 хв. За відмінностями у послідовності цього гена судять про спорідненість у багатьох таксонах тварин.
Матеріал надсилався до компанії «Macrogen Europe» у Амстердамі, де проводилось секвенування гену CO.
Біоінформаційні дослідження проводились безпосередньо мною. Для початку знаходили споріднені види шляхом пошуку у Генбанку NCBI методом BLAST [15]. Цей метод підбирає найбільш подібні послідовності з наявних у базі даних, що дало нам певну кількість видів, які мають найбільшу спорідненість з досліджуваним об’єктом. Потім всі ці послідовності завантажувались у програму «BioEdit» [14] для вирівнювання (зіставлення) генів та редагування (у послідовностях з Генбанку був присутній один повний мітохондріальний геном, а не тільки один ген). Вирівнювання потрібно для коректної роботи програм для будування філогенетичних дерев та загалом для оцінки змін у послідовностях у різних видах.
Далі вирівняні послідовності завантажувались у програму «MEGA-X» [13], у ній я будував філогенетичні дерева методами Neighbor Joining [10] та Maximum Likelihood з еволюційною моделлю під назвою «Tamura-3-parameter model» [12] та 500 реплікаціями бутстрепінгу (кількість свідомо поданих несправжніх даних, які програма аналізує для вирахування довірчої оцінки компонентів дерева) [11]. Філогенетичні дерева відображають спорідненість видів та, фактично, шляхи еволюції.
Сутність першого методу полягає у порівнянні генетичних відстаней. На вході є набір послідовностей, які спочатку у вигляді дерева подаються як несистематизована «зірочка». Далі, програма порівнює генетичні відстані (генетична відстань – величина, що є різницею між послідовностями та є значенням кількості відмінних нуклеотидів) всіх гілок та обирає поміж них гілки з найменшою відстанню, які об’єднуються в одну групу, після чого всі інші гілки знов порівнюються (вже з урахуванням цієї групи), і так до кінця, поки не буде опрацьований увесь масив даних.
Сутність же другого методу полягає у переборі всіх можливих варіантів дерев з пошуком тих, які будуть найбільше відповідати вхідним даним у вигляді вирівняних нуклеотидних послідовностей. Далі філогенетичні дерева редагували у програмі «FigTree» та «Inkscape», у яких отримані дерева обробляли для надання їм більш якісного та наочного вигляду. Потім дерева аналізували, і з них ми робили відповідні висновки.
Також ми провели морфологічні дослідження. Дослідження морфології проводили на кафедрі зоології та екології тварин Харківського національного університету імені В.Н. Каразіна за допомогою мікроскопа Konus Crystal-45.
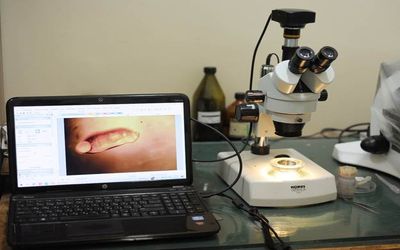
Екземпляри розміщували у спиртовому розчині, виставляли у потрібне положення за допомогою пінцета, після чого, регулюючи збільшення й фокусну відстань мікроскопа, досягали максимальної різкості. Далі за допомогою камери для мікрофотозйомки, яка приєднувалась до мікроскопа, зображення виводилось на монітор комп’ютера, де за допомогою спеціального додатку робили корекцію та, власне, фото. Потім фотографії аналізували та обробляли у редакторах для надання їм більш наочного вигляду.
Філогенетичне дослідження
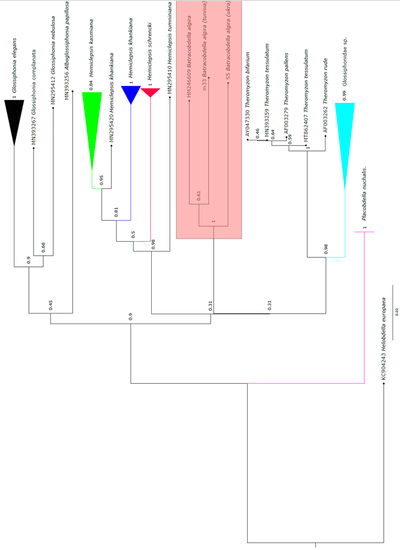
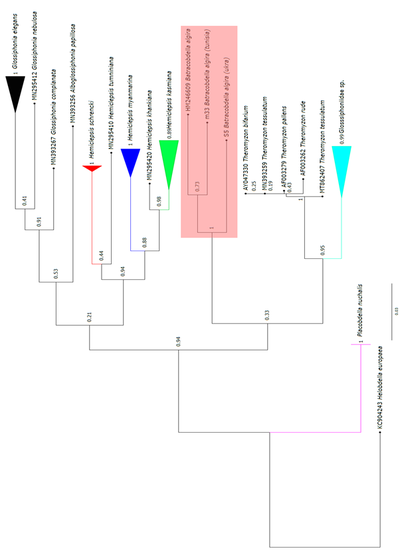
Ми проаналізували обидва філогенетичні дерева, які представлені на рис. 3.1, 3.2. По-перше, ми бачимо, що дерева дуже схожі, що свідчить про високий рівень достовірності результатів їх побудови. З дерева, побудованого методом Neighbor Joining, ми бачимо, що різниця помітна для всіх трьох видів.
Досить висока підтримка бутстрепінгу каже про те, що і іспанський екземпляр (з Генбанку) може належати до окремого виду, а от відмінність українського екземпляра є дуже вірогідною, що каже про те, що ця п’явка точно вже не належить до виду жабоп’явка алжирська. Такі самі результати показав й метод Maximum Likelihood: є певна різниця між туніським та іспанським екземплярами, а от стверджувати про те, що кримська популяція не є жабоп’явкою алжирською, можна майже зі 100% ймовірністю.
Морфологічне дослідження
Результати морфологічного дослідження показали, що, по-перше, ці п’явки дійсно належать до цього роду, бо у фіксованому стані специфічно вигинають черевну поверхню у вигляді «човника». По-друге, кримський екземпляр відповідає тому, що було описано про B. algiraраніше і є дуже схожим з ним за зовнішньою будовою, однак суттєво відрізняється вираженість, розмір і кількість сосочків – у кримського екземпляра їх значно менше, вони менші за розміром та вони значно гірше помітні у порівнянні з туніським екземпляром.
Крім того, є різниця у формі заднього присоска – у кримського екземпляра видна окантовка, у той час, як у туніського екземпляра вона є більш пласкою. Окрім цього, у туніського екземпляра є набагато помітніша пігментація в порівнянні з кримським, але це може бути результатом різних методик фіксації. Тож, казати саме про відмінності у пігментації передчасно. Втім, як ми бачимо і як я описав вище, певна різниця все-таки є, що ще раз підтверджує різницю цих видів. Мікрофотографії морфологічних досліджень представлені у додатках №1-№14.
Виходячи з широкого, як вважалося, ареалу B. algira, ми припускаємо, наступний сценарій видоутворення: спочатку B. algira(чи її пращур) дійсно займала великий ареал (дивитись додаток №21), але взнаки давалась географічна ізоляція багатьох популяцій (наприклад у Криму). Поступово кожна популяція в різних ареалах почала незалежно змінюватись завдяки природному добору, а різність умов спричиняла дивергенцію.
Таким чином, за певний час, різні популяції накопили таку кількість змін (причому різні популяції еволюціонували з різною швидкістю), що вони тепер вже не належать до виду B. algira. Ми припускаємо, що кримська популяція еволюціонувала за вищеописаним сценарієм (фактично за алопатричним типом видоутворення) та є новим видом, а враховуючи ізольованість Криму, найімовірніше, ендемічним (втім, потрібні додаткові дослідження для доведення ендемічності).
Стан кримської популяції
Наразі для цього виду в Криму існує небезпека. По-перше, вже зараз ця популяція під назвою жабоп’явка алжирська занесена до Червоної книги України зі статусом «вразлива» [6]. Тепер же ми знаємо, що цей статус треба переглянути, враховуючи ендемічність нового виду.
По-друге, як ми знаємо, у Криму відбувається гуманітарна й екологічна катастрофа, пов’язана з нестачею прісної води [7], яка може сильно зашкодити популяції цього виду, адже вони розповсюджені лише у прісних водоймах. Задля збереження цілісності популяції необхідно покращити загальний екологічний стан у регіоні та відновити кількість водойм, де ці п’явки живуть.
Висновки
У всіх літературних джерелах (в тому числі й такому важливому, як «Червона книга України») популяція кримських жабоп’явок згадується як Batracobdella algira. Втім, відсутні будь-які молекулярно-генетичні дослідження і морфологічні порівняння для точного встановлення її таксономічного статусу
За результатами філогенетичного аналізу, кримський екземпляр є окремим видом, відмінним від B. algira, оскільки різниця у генетичних послідовностях є досить великою і обидва методи побудови філогенетичних дерев поміщали його в окрему гілку.
Іспанський та туніський екземпляри також можуть належати до різних видів, оскільки їхня еволюція могла йти різними шляхами, що добре помітно на філогенетичному дереві.
Морфологічні дослідження підтверджують висунуту нами гіпотезу – екземпляри досить суттєво відрізняються принаймні сосочками. Крім того, є кілька інших відмінностей, які також можуть свідчити на користь самостійності цих двох видів
В результаті дослідницької роботи (проєкта) з біології у 11 класі ми припускаємо, що кримська популяція жабоп’явки алжирської є новим (найімовірніше ендемічним) видом, який відділився від спільного пращура з B. algira за алопатричним типом видоутворення. Тож тепер ця популяція потребує опису як нового виду.
Список використаних дежерел та літератури
- Lucinda P. Lawson, John M. Bates, Michele Menegon, Simon P. Loader Divergence at the edges: peripatric isolation in the montane spiny throated reed frog complex. BMC journal. 2015. 1 лип. (№128)
- И. С. Даревский, В. В. Гречко, Л. А. Куприянова Ящерицы, размножающиеся без самцов. Журнал Природа. 2000. №9
- Roman Yulkievich The rate test of specifications: estimating likelihood of non-allopatric speciation from reproductive isolation rates of Drosophila. Pubmed journal. 2014.
- Schuichi Shikano, Leo S. Luckinbill, Yasushi Kuribara Changes of Traits in a Bacterial Populations Associated with Protozoal Predation. Microbial Ecology. 1990. 18 трав. (№20). С 75-84
- Фауна України [Текст] : в сорока томах / Академія наук Української РСР, Інститут зоології ім. І.І. Шмальгаузена. – Київ : Наукова думка, 1956 – Том 30 : П’явки : зовнішня і внутрішня будова, екологія, систематика, поширення та практичне значення п’явок / Є. І. Лукін. – Київ : Видавництво АН Української РСР, 1962. – 93 с.
- Червона книга України: Жабоп’явка алжирська Bactracobdella algira (Moquin-Tandon, 1846)
- Перший український інформаційний телеканал «5 канал»: В окупованому Криму назріла екологічна катастрофа (дата звернення: 2.12.2020)
- Andrii Khomenko, Serge Utevsky, Andriy Utevsky, Peter Trontelj Unrecognized diversity of Trocheta species (Hirudinea: Erpobdellidae): resolving a century-old taxonomic problem in Crimean leeches. Systematics and Biodiversity journal. 2020. 23 квіт. С 1-13
- Ben Ahmed R., Ropelewska E., Bielecki A., Cichocka J. Bactracobdella algira Moquin-Tandon 1846 (Hirudinida: Glossiphoniidae)—morphometric analysis and internal morphology. Europe PMC journal. 2009. 1 січ. С 353-358
- Saitou N. and Nei M. (1987). The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 4:406-425.
- Felsenstein J. (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39:783-791.
- Tamura K. (1992). Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Molecular Biology and Evolution 9:678-687.
- Kumar S., Stecher G., Li M., Knyaz C., and Tamura K. (2018). MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Molecular Biology and Evolution 35:1547-1549.
- Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41:95-98.
- S. Altschul, W. Gish, W. Miller, E. Myers, and D. Lipman. Basic local alignment search tool. Journal of Molecular Biology, 215(3):403-410, October 5, 1990.
Додатки


Нові проєкти і роботи




Навчальні програми



Банер сайту

Код банера:
