Дослідницькі роботи і проєкти
Вплив щільності популяції плодової мушки дрозофіли на плодючість

У процесі написання науково-дослідницької роботи (проєкта) з біології про вплив щільності популяції плодової мушки дрозофіли на плодючість самок учениця 11 (10) класу вивчає поняття популяції та її характеристик, розглядає популяції комах Drosophila melanogaster. Також в міні-проєкті розглядаються фактори, які впливають на популяцію плодової мушки дрозофіли, на розмноження комах і їх плодючість.
Докладніше про роботу:
Протягом дослідницької роботи ученицею 11 класу було проведене дослідження з біології, за допомогою якого виявлено вплив щільності популяції плодової мушки дрозофіли на її плодючість і середню вагу нащадків, розглянут статевий диморфізм плодової мушки та цикл розвитку плодової мушки - Drosophila melanogaster. В дослідницькій роботі над проєктом досліджується цикл розвитку плодової мушки, а також автором проводиться експеримент з визначення впливу щільності популяції дрозофіли на плодючість самок.
В запропонованому творчому навчальному проєкті з біології про вплив щільності популяції плодової мушки дрозофіли на плодючість її самок з’ясовується, що таке плодючість і які чинники впливають на плодючість комах Drosophila melanogaster, досліджується вплив щільності популяції плодової мушки на плодючість самок і середню вагу нащадків.
Зміст
Вступ
1. Поняття про популяцію та її основні характеристики.
2. Популяції комах і фактори які на них впливають.
3. Розмноження комах і їх плодючість.
4. Вплив щільності популяції плодової мушки на її плодючість і середню вагу нащадків.
4.1. Статевий диморфізм плодової мушки.
4.2. Цикл розвитку плодової мушки.
4.3. Експеримент з визначення впливу щільності популяції плодової мушки на плодючість самок.
Висновки
Список використаних джерел.
Вступ
В природі живі організми існують не у вигляді окремих особин, вони мешкають групами, займаючи окрему територію – ареал. Такі групи особин одного виду зі спільним генофондом, спільною морфологією і єдиним життєвим циклом називають популяцією. Кожен вид являє собою складний комплекс внутрішньовидових груп, які охоплюють в своєму складі особин зі специфічними рисами будови, фізіології та поведінки.
Вивчення популяцій важливе для прогнозування змін, які в них відбуваються і їх регулювання. Наприклад, при заготівлі деревини важливо знати швидкість відновлення лісу, щоб правильно планувати інтенсивність рубок. Аналогічна ситуація з популяціями тварин, які людина використовує як харчову або хутряну сировину. З медико-санітарної точки зору велике практичне значення має вивчення популяцій дрібних гризунів, які є носіями небезпечної для людини хвороби – чуми, або розповсюдження щурів, які є носіями і розповсюджувачами лептоспірозу [1].
Вже багато десятиліть людина вивчає вплив різних факторів і чинників на популяції різних видів тварин. Розуміння основних закономірностей, за якими відбувається збільшення або зменшення чисельності популяції, дає людині можливість впливати на шкідників біологічними методами, охороняти рідкісні види тварин або вирощувати їх в штучних умовах.
Одним із перспективних напрямків на сьогодні є вирощування комах в неволі. Останнім часом інтерес до масового розведення комах як у нас у країні, так і за кордоном, різко зріс. Обумовлено це тими можливостями, які відкриває розведення комах для вирішення актуальних завдань прикладної ентомології [3].
Так, на сьогодні актуальним є розведення господарсько-корисних видів комах – продуцентів сировини і продуктів харчування, також багато культур комах використовуються в сільському господарстві для захисту рослин, тварин і людини від шкідливих членистоногих. Ще одним аспектом цього питання є розведення комах, чисельність яких у природі постійно зменшується. Серед таких видів можна виділити об’єкти естетичної насолоди людини (в основному це яскраво забарвлені тропічні та субтропічні види комах) та види, що підлягають охороні [5].
Актуальність роботи: Одним з ефективних методів відновлення популяцій зникаючих видів комах є розробка методів їх лабораторного культивування. При розробці і застосуванні цих методів важливо розуміти всі фактори і чинники які впливають на розвиток популяції комах в штучних умовах. Саме тому, ми вирішили дослідити вплив такого фактору як щільність популяції комах під час їх штучного розведення.
Об’єктом нашого дослідження ми обрали плодову мушку Drosophila melanogaster.
Мета роботи: дослідити вплив щільності популяції плодової мушки на плодючість самок і середню вагу нащадків.
Для досягнення поставленої мети необхідно було вирішити такі завдання:
- Визначити основні характеристики популяції комах і фактори, які на них впливають.
- З’ясувати, що таке плодючість і які чинники впливають на плодючість комах.
- Дослідити вплив щільності популяції плодової мушки на плодючість самок і середню вагу нащадків.
Поняття про популяцію та її основні характеристики
Вивчення популяцій, їх взаємовідносин і динаміки є одним із основних завдань екології [2]. Термін «популяція» вперше був введений датським вченим Йогансеном у 1903 році для визначення «природної суміші особин одного і того ж виду, неоднорідної в генетичному відношенні».
За визначенням С.С. Шварца (1980) «популяція – це елементарне угрупування організмів певного виду, які мають всі необхідні умови для підтримання своєї чисельності певний проміжок часу в певних умовах середовища, що постійно змінюються».
На сьогодні під поняттям популяція розуміють сукупність особин певного виду, які тривалий час (багато поколінь) живуть на певній території і вільно схрещуються між собою. Водночас певна популяція під впливом якихось чинників відокремлена від територій інших аналогічних популяцій. Популяція сама по собі може підтримувати свою чисельність необмежений час [4].
З визначення випливає, що можна запропонувати певну ієрархію популяцій:
- елементарна, або локальна — означає найменшу сукупність, яка живе на невеликій однорідній ділянці (у певному лісі, ставку);
- екологічна — складається з елементарних популяцій і займає вже значно більшу територію;
- географічна — охоплює велику сукупність особин одного виду на великій території з приблизно однаковими умовами. У межах такої популяції відмінність особин невелика, але вона помітна, якщо їх порівнювати з представниками іншої географічної популяції.
Як кожна складна система популяція має чимало характеристик, найголовнішими з них є:
- чисельність;
- густота розселення;
- генетична структура;
- дані про імовірність виживання;
- тип просторового розподілу особин;
- розподіл особин за віком (демографічна піраміда);
- спосіб розмноження і плодючість;
- соціальна організація.
Порівняно легко підрахувати чисельність популяції великих тварин на відкритих просторах (оленів у тундрі, дельфінів у морі), але складніше це зробити для невеликих чи мікроскопічних істот, які до того ж можуть вести нічне чи підземне (підводне) життя. В цих умовах бажано виміряти середню густоту заселення і визначити тип та величину площі розселення.
Розрізняють кілька типів організації розташування особин у популяції:
- випадкове (хаотичне) і впорядковане,
- рівномірне і групове (нерівномірне).
Популяції характеризують параметрами:
- Чисельність – загальна кількість особин в популяції.
- Запас біомаси популяції.
- Щільність – кількість особин на одиницю території (або об’єму простору).
- Народжуваність – кількість нових особин за певний проміжок часу на одну особину.
- Смертність – кількість померлих особин за певний проміжок часу.
- Ріст популяції – співвідношення народжуваності та смертності.
Залежно від певних характеристик популяції її структуру підрозділяють за показниками:
1. Генетична структура властива популяціям, які мають дві або більше генетичних форм, що обумовлює генетичний поліморфізм популяції і збільшує її стійкість.
2. Статева структура популяції – співвідношення особин різної статі. При генетичному контролі це співвідношення дорівнює 50×50. При впливі природного середовища воно змінюється в бік більшої смертності самців.
3. Вікова структура властива природнім угрупованням тварин та рослин, якщо вони не однолітні. В популяціях культурних рослин, в яких завдяки одночасному посіву вік особин однаковий, вікова структура не має значення.
4. Розмірна структура популяцій визначає відмінності особин за розмірами.
5. Просторова структура популяції визначає характер розміщення особин на території ареалу.
Популяції комах і фактори які на них впливають
Комахи як складова частина біогеоценозів суходолу та водойм зазнають впливу різноманітних факторів навколишнього середовища, у свою чергу впливаючи на нього [6].
Абіотичні фактори. У зв'язку з тим що комахи – пойкілотермні тварини, їх поведінка, життєдіяльність, тривалість розвитку, а також популяційна динаміка часто визначаються температурними умовами середовища, що набувають значення головного екологічного фактора. Активна життєдіяльність комах можлива лише у межах певного діапазону температур – від +10°С до +45°С (при оптимумі від +25° до +27°С).
Поза цією температурною зоною активності комахи впадають у стан або холодового, або теплового заціпеніння, з якого вони можуть вийти при поверненні температури в межі зони активності. Проте сильне підвищення чи зниження температури може спричинити загибель комах. Окремі види здатні витримувати значні зниження або підвищення температури. Наприклад, личинки веснянок і комарів, що населяють водойми тундри, активні при 0°С. Личинки стеблового метелика у стані діапаузи можуть витримувати охолодження до -25°С, озимої совки – до -11°С. Одночасно личинки деяких видів розвиваються в гарячих джерелах із температурою до +70°С (африканський комарик) [6, 7].
Комахи чутливі до вмісту вологи в середовищі. Вологолюбні (наприклад, цвіркун, стебловий метелик) заселяють долини річок та інші зволожені місця, де відносна вологість повітря становить 80-100% . Водночас є види, які пристосувалися жити при посушливому кліматі пустель і напівпустель (пустельна сарана, жуки-чорнотілки, жужелиці, мурашки, клопи). Переважна більшість видів комах нашої фауни потребує помірної вологості (у межах 50-80%). Важливе значення має і вологість ґрунту. Відомо, що колорадські жуки навесні із сухого ґрунту виходять пізніше або не виходять зовсім доти, поки не буде опадів.
Багато комах п'є воду. Волога впливає на комах разом із температурою, тому сумісну дію цих факторів розглядають як гігротермічний режим. Так, для розвитку личинок колорадського жука оптимальним є такий гігротермічний режим: відносна вологість – 60-70%, температура – 24°-25°С. Якщо личинки розвиваються у дощову холодну погоду, вихід їх із яєць становить лише 25%.
Світло значною мірою впливає на фізичні й хімічні процеси в організмі комах, обмін речовин, поведінку, розвиток, розселення в біотопі [6, 7].
У розвитку комах існує стан тимчасового фізіологічного спокою – діапауза. Вона характеризується різким зниженням обміну речовин та припиненням формотворних процесів. Діапауза виникла в процесі еволюції як одне із пристосувань до перенесення несприятливих умов навколишнього середовища взимку, а в посушливому кліматі – улітку. Діапауза перебуває під контролем тих факторів середовища, які мають правильну сезонну періодичність.
Такими є довжина світлового дня (фотоперіод), температура і вологість повітря, біохімічний стан рослин. Серед цих факторів вирішальне значення має фотоперіод – асторономічно точний сигналізатор про наступну зміну життєвих умов. Проте було помічено, що висока температура восени навіть при короткому дні гальмує настання діапаузи у деяких комах, зокрема, у колорадського жука.
Для комах характерний добовий ритм активності. Є комахи, активні вдень (саранові, денні метелики, перетинчастокрилі, мухи та ін.). У присмеркових та нічних комах підвищена активність спостерігається ввечері та вночі (цвіркуни, бражники, комарі, совки та ін.).
Вітер як фактор середовища впливає на розселення комах. Багато дрібних комах (попелиці, мошки, деякі метелики) пасивно переносяться потоками повітря. Вертикальні повітряні потоки піднімають комах на висоту до 1-2 тис. м.
 Біотичні фактори. Важливу роль у житті комах відіграють їхні взаємозв'язки із рослинами та іншими тваринами, насамперед, харчові, або трофічні. За способом живлення серед комах є фітофаги (саранові, довгоносики, попелиці, короїди, вусачі, листоїди та ін.), хижаки (жужелиці, кокцинеліди, бабки, богомоли, ктирі та ін.), паразити (їздці, мухи-тахіни, ґедзі, пухоїди, воші, блохи), сапрофаги (личинки деяких жуків, двокрилих), некрофаги (жуки-мертвоїди, личинки мух, мурашки), копрофаги (жуки-гнойовики, мухи, терміти), всеїдні (таргани).
Біотичні фактори. Важливу роль у житті комах відіграють їхні взаємозв'язки із рослинами та іншими тваринами, насамперед, харчові, або трофічні. За способом живлення серед комах є фітофаги (саранові, довгоносики, попелиці, короїди, вусачі, листоїди та ін.), хижаки (жужелиці, кокцинеліди, бабки, богомоли, ктирі та ін.), паразити (їздці, мухи-тахіни, ґедзі, пухоїди, воші, блохи), сапрофаги (личинки деяких жуків, двокрилих), некрофаги (жуки-мертвоїди, личинки мух, мурашки), копрофаги (жуки-гнойовики, мухи, терміти), всеїдні (таргани).
Серед комах є й такі, що живляться роговою речовиною – кератофаги (гр. keras - «ріг»): представники родин шкіроїди (Dermestidae), облудники (Ptinidae), пухоїди (Mallophagaа); а також воском (вогнівка воскова). Якість корму впливає на обмін речовин, ріст і розвиток комах. Наприклад, при живленні картоплею личинки колорадського жука розвиваються протягом 13-19 днів, на синіх баклажанах – 13-21, помідорах – 25-32, блекоті – 30-48 днів.
Живлення – вирішальний фактор зміни чисельності популяції виду. Ним зумовлюється плодючість і виживання потомства. Так, живлення гусениць непарного шовкопряда різними рослинами позначається на плодючості самок: самки з гусениць, що живилися листям дуба і граба, відкладають 500-600 яєць, яблуні – 384, берези – 286. Розвиток гусениць триває: на дубі – 40 днів, грабі – 44, яблуні – 46, глоді – 55.
У біоценозах комахи перебувають у складних взаємозв'язках з іншими тваринами. Серед комах є екто- та ендопаразити майже всіх класів тварин, а також хижаки що живляться червами, молюсками, членистоногими, мальками риб, пуголовками. Хижаків та паразитів, що живуть за рахунок інших комах, об'єднують у групу ентомофагів.
Вони регулюють чисельність комах-шкідників, перешкоджаючи їхньому масовому розмноженню. До ентомофагів належать жужелиці, сонечка, мурашки, яйцеїди, мухи-тахіни та ін. У свою чергу комахи – незамінна їжа хребетних усіх класів (особливо птахів, плазунів, земноводних та риб).
У процесі еволюції в комах з'явився ряд пристосувань до виживання: захисне та застережне забарвлення, здатність виділяти отруйні речовини, мімікрія та ін. [6,7].
Антропогенні фактори. Діяльність людини (освоєння під сільськогосподарські культури степів та інших цілинних земель, вирубка і викорчовування лісів, осушення боліт, зрошення земель, випасання худоби, технічне і побутове забруднення довкілля та ін.) впливає на склад ентомофауни та чисельність комах. Одні види при цьому витісняються і зникають, інші, навпаки, інтенсивно розмножуються.
У культурних біоценозах, зокрема, в агроценозах, де переважає монокультура рослин (пшеничне, картопляне поле та ін.), створюються умови для процвітання невеликої кількості видів комах, які живляться певним видом рослин. Достатня кількість їжі та відсутність природних ворогів сприяють їх масовому розмноженню, різкому збільшенню чисельності. Так людина сама створює умови для появи комах-шкідників.
Розвиток транспорту посилює і прискорює розселення комах далеко за межі їхнього природного ареалу. Значну роль відіграло транспортування вантажів у розселенні виноградної філоксери, колорадського жука, американського білого метелика, немирного шовкопряда та інших небезпечних шкідників.
З комахами-шкідниками людина проводить боротьбу. На основі глибокого вивчення екології комах розроблено методи прогнозування та регулювання чисельності шкідників. Розрізняють 5 основних груп: хімічні, біологічні, агротехнічні, механічні ти карантинні методи [6,7].
Розмноження комах і їх плодючість
Комахи – переважно роздільностатеві тварини. Вони розмножуються лише статевим способом. Запліднення яєць у комах відбувається тільки всередині організму самки. У деяких випадках розмноження комах може відбуватися без запліднення. Такий спосіб розмноження називається партеногенезом [6].
Комахам властивий складний життєвий цикл. Процес постембріонального розвитку відбувається в них із метаморфозом – глибоким перетворенням організму, що проявляється в різкій зміні будови й способу життя. У всіх представників цього класу спочатку з яйця з’являється личинка, яка згодом перетворюється на дорослу комаху.
Неповний метаморфоз спостерігається у комах простої організації: сарани, тарганів, бабок та одноденок. В їхньому розвитку виділяють три стадії: яйце – личинка – доросла комаха. За умови неповного метаморфозу з яйця виходить маленька безкрила личинка, яка дуже схожа на дорослу особину і веде подібний спосіб життя.
Від дорослої комахи ця личинка – німфа (від грец. німфе – дівчина) відрізняється головним чином розмірами і відсутністю крил. Підростаючи, личинка линяє – виповзає зі старого хітинового покриву в новому, просторішому вбранні. Після кожного линяння німфа стає дедалі більше схожою на дорослу комаху. У процесі розвитку личинка линяє 5-6 раз. Дорослі комахи більше не ростуть і не линяють.
Повний метаморфоз властивий високоорганізованим комахам: жукам, метеликам, мухам, комарам, бджолам та блохам. В їхньому розвитку виділяють чотири стадії: яйце – личинка – лялечка – доросла комаха.
Личинки комах з повним перетворенням істотно відрізняються від дорослих особин за певними ознаками будови й способу життя:
- форма тіла личинок переважно червоподібна;
- вони не мають крил;
- органи чуття в личинок недорозвинені, відсутні фасеткові очі, наявні лише прості вічка. Личинка мухи навіть не має голови, тому її очі розташовані на грудях;
- часто спостерігається відмінність у кількості і будові кінцівок. Так, личинка метелика – гусениця – крім трьох пар справжніх ходильних ніг, має ще й черевні ніжки з присосками, які ніколи не залишаються в дорослого метелика. Личинка мухи, навпаки, не має ніг;
- личинки часто живуть в іншому середовищі і мають свій особливий спосіб живлення. Так, личинка комара живе у воді і живиться дрібнішими тваринами й водоростями, а дорослий комар – літаюча комаха, яка живиться соком рослин і кров’ю хребетних тварин.
Лялечка комахи не живиться і звичайно майже нерухома. Усередині лялечки відбувається складна внутрішня перебудова, за якої більша частина органів личинки руйнується, наче розчинюється, а натомість із скупчень спеціальних клітин формуються органи дорослої комахи.
Більшості комах властивий розвиток із повним метаморфозом. Завдяки відмінностям у будові, способі життя, живленні, а також у середовищі існування комахи з повним метаморфозом краще пристосувалися до природного середовища, ніж комахи з неповним метаморфозом. Тому перші складають понад 90 % усіх видів комах.
Плодючість комах хоч і значна, але не є постійною величиною. Показник визначається двома факторами: спадковими властивостями виду (будова і величина яєчників) – потенціалом розмноження і впливом навколишнього середовища. За сприятливих зовнішніх умов потенціал розмноження реалізується найбільшою мірою.
Так, самка бавовникової совки за своє життя може відкласти близько 2700, озима совка – 1800, лучний метелик – 800, гессенська муха – 500, п’явиця червоногруда – 300, хлібний пильщик – 50 яєць. В окремих випадках плодючість може сягати разючих розмірів, особливо у суспільних комах. Наприклад, матка медоносної бджоли відкладає до 3 тис. яєць за день, а у термітів – навіть до 30 тис. Але потенціал розмноження не завжди реалізується повністю, плодючість може бути істотно знижена під впливом несприятливих умов середовища – погоди, живлення тощо [6, 7].
Вплив щільності популяції плодової мушки на її плодючість і середню вагу нащадків
Статевий диморфізм плодової мушки
Об’єктом наших досліджень ми обрали плодову мушку Drosophila melanogaster.
Drosophila melanogaster, інакше плодова, чи оцтова муха – маленька, близько 3 мм завдовжки, комаха сімейства Drosophiliadae з ряду Diptera. Цих мушок можна спостерігати близько зіпсованих фруктів.
Дрозофіла стала однією з цінних для біологічних досліджень організмів, зокрема, у сфері генетики. Її використовують, як модельний організм щодо різноманітних досліджень вже майже сто років, і сьогодні багато вчених продовжують працювати над різними проблемами дрозофіли.
Дрозофіла – маленька тварина з коротким життєвим циклом тривалістю всього лише близько двох тижнів, вона недорога для розмноження і виведення навіть великої кількості особин. Дрозофіла спочатку використовувалася для генетичних досліджень, наприклад, з вивчення закономірностей різних типів спадковості. Короткий період розвитку від яйця до імаго, виняткове різноманіття мутаційних рас із властиво чітким фенотиповим проявом, мала кількість хромосом й інших важливих переваг роблять її незамінною для практичного вивчення основних закономірностей спадковості.
Морфологічно, самки і самці відрізняються один від одного цілою низкою ознак. Самки трохи більші за самців. Черевце у самки дещо округле, із загостреним кінцем; у самця воно циліндричне, з притупленим кінцем (рис. 1). Також самця можна легко відрізнити від самки по кількох останніх сегментах, які у нього суцільно пігментовані.
Верхні хітинові щитки на грудях у комах називаються тергітами (вони разом зі стернітами і плейрітами беруть участь в русі крил). Статевий диморфізм у дрозофіли виявляється в тому, що у самки є вісім добре розвинених тергітів, а у самця – шість, причому шостий і восьмий тергіти злиті, а восьмий увійшов до складу статевого апарату. Стерніти – такі ж хітинові пластинки з черевного боку (рис.1). У самки їх також на один більше, ніж у самця, і у представників кожної статі не розвинені перший, другий, сьомий і восьмий стерніти.
До числа вторинно-статевих ознак у самця відносяться статеві гребінці,що представляють собою міцні хітинові щетинки на перший членик лапки передніх ніг. У самки подібні утворення відсутні.


Цикл розвитку плодової мушки
Яйця дрозофіли декілька витягнуті, близько 0,5 мм у довжину. У свіжих культурах вони легко помітні, будучи відкладеними на живильному середовищі, здебільшого поблизу від стінок пробірки, де міститься менше вологи. Яйце захищене двома зовнішніми оболонками, різними за походженням. Запліднення яйця відбувається в момент проходження його через верхній відділ піхви. У нормальних умовах ембріональний розвиток протікає поза тіла матері при температурі 270С близько 20 годин.
Вилуплення личинки з яйця і початок постембріонального розвитку пов'язано з посиленим живленням і зростанням. Надлишок їжі на цій стадії життя особи має велике значення: у значній мірі він визначає не тільки розміри мухи, але і її життєздатність. Перший час після вилуплення личинки залишаються на поверхні середовища. Потім вони ідуть у глиб її і залишаються там до моменту окуклення.
Окуклення починається з того, що личинки залишають середу, перестають живитися і якийсь час жваво повзають по стінках пробірки. Потім вони стають нерухомими, значно скорочуються в довжину і набувають характерну для лялечки діжкоподібну форму. Період лялечкового перетворення дорівнює в середньому 4 доби.
Після закінчення третьої доби через покрив лялечки стають помітними обриси очей, в яких на той час виробляється жовтуватий пігмент. За кілька годин до вилуплення добре видно крила, очі до цього часу набувають яскраво-червоного кольору (рис. 2).
 Рис. 2 Цикл розвитку дрозофіли |
Зазвичай мухи виходять з лялечки рано вранці. Мухи, які щойно вилупилися, мають довге жовтувате тіло, майже позбавлене пігменту, короткі, ще не розправлені крила. Через 8 годин самки вже готові до запліднення. |
Експеримент з визначення впливу щільності популяції плодової мушки на плодючість самок
Для вирощування мух ми приготували спеціальне поживне середовище: на 100 г води додали 6 г агар-агару, 5 г цукру, 5 г манної крупи, 7 г дріжджів. Після приготування середовище розлили у спеціальні пробірки з широким дном по 5 мл в кожну. Поверхню середовища змастили суспензією живих дріжджів, якими живляться дрозофіли в імагінальному стані. Середовище є кормом для личинок, заварені дріжджі є для них джерелом протеїнів.
Для розсадки мух ми використовували спеціальний столик, дзеркальце (на яке висипали мух), пензлик і морилку. Мух присипляли ацетоном.
В 5 різних пробірок ми розсадили різну кількість пар (родин) самців і самок: 2, 5, 10, 15, 20 пар (табл. 1). Пробірки закрили корками і помістили їх у термостат з температурою 23-25 0С на 12-14 діб. На 12 день почався вихід мух. Результати експерименту підраховували на 14 день.
Таблиця 1. Експеримент по плодючості дрозофіли
| № пробірки | Кількість родин | Кількість нащадків (шт.) | Кількість нащадків (шт.) | Кількість нащадків (шт.) | Плодовитість 1 самки1 шт |
Середня вага самця2 мг |
Середня вага самки3 мг |
| Всього | ♂ | ♀ | |||||
| 1 | 2 | 45 | 29 | 16 | 22,5 | 1,5 | 1,62 |
| 2 | 5 | 36 | 21 | 15 | 7,2 | 1,41 | 1,52 |
| 3 | 10 | 55 | 25 | 30 | 5,5 | 1,35 | 1,45 |
| 4 | 15 | 47 | 25 | 22 | 3,1 | 1,22 | 1,32 |
| 5 | 20 | 51 | 27 | 24 | 2,5 | 1,15 | 1,21 |
1 – nнащ = n всіх нащадків / nсамок; 2 – mсер♂ = m10♂ / 10; 3 – mсер♀ = m10♀ / 10
Після повного виходу мух, ми окремо дослідили результати в кожній пробірці. Для цього мух з кожної пробірки почергово заморювали, висипали на дзеркальце, перераховували загальну кількість нащадків. Вираховували плодючість однієї самки за формулою: nнащ = n всіх нащадків / nсамок (табл. 1).
Потім сортували окремо самців і самок, підраховували їх кількість. З кожної пробірки відбирали по 10 особин однієї статі, зважували на торсійних вагах і вираховували середню масу однієї особини за формулою: mсер♀ = m10♀ / 10, так само і для самців (табл. 1).
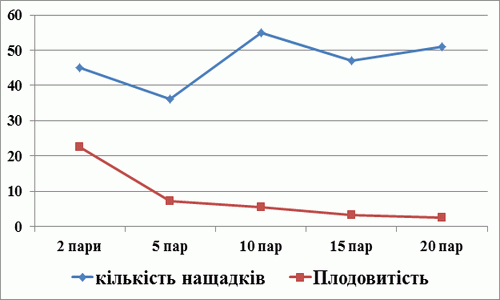
У результаті проведених досліджень установлено, що найбільша індивідуальна плодючість самок спостерігається в популяції із 2-х родин (пробірка №1). Плодовитість однієї самки склала 22,5 нащадки (табл. 1). Оцінюючи вплив щільності популяції плодової мушки на загальну кількість нащадків (рис. 3), ми встановили, що найбільша кількість нащадків була у пробірці №3, в якій було посаджено 10 сімей.
Але аналіз індивідуальної плодючості самок показав, що із збільшенням щільності популяції плодової мушки в пробірці індивідуальна плодючість самок різко зменшується (рис. 3). При достатній кількості поживного середовища індивідуальна плодючість самок плодової мушки складає 20-22 нащадки, що ми і спостерігали у пробірці №1.
Із збільшенням щільності мух у пробірці настає дефіцит поживного середовища, що і є основним чинником впливу на індивідуальну плодючість самок. Тому, при вирощуванні плодової мушки в лабораторних умовах, оптимальною кількістю на 5 мл поживного середовища є 2 пари мух (2 сім’ї).
Результати досліду по впливу щільності популяції плодової мушки на середню масу нащадків показали, що із збільшенням щільності маса як самців так і самок зменшується (рис. 4). Дефіцит поживних речовин впливає на нормальний розвиток личинок, маса нащадків значно зменшується в наслідок збільшення кількості сімей у пробірках.
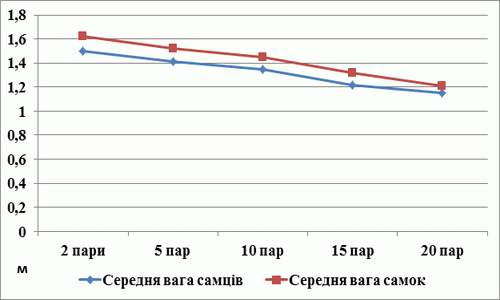
Таким чином, у результаті проведених досліджень установлено, що збільшення щільності популяції плодової мушки впливає на індивідуальну плодючість самок і на середню вагу їх нащадків. При розведенні комах у лабораторних умовах важливо правильно підбирати поживне середовище і його кількість, враховувати оптимальну щільність популяції при закладці досліду, бо це може вплинути на індивідуальну плодючість самок і якість отриманих нащадків.
Висновки
- На розмноження і розвиток комах впливає ряд абіотичних і біотичних факторів, а також антропогенний чинник.
- Встановлено, що збільшення щільності популяції плодової мушки в лабораторних умовах знижує індивідуальну плодючість самок.
- В дослідницькій роботі над проєктом з’ясовано, що збільшення щільності популяції плодової мушки впливає на середню масу нащадків. Зі збільшенням щільності середня маса як самців так і самок зменшується.
- При розведенні комах в лабораторних умовах потрібно враховувати такий чинник як щільність популяції та кількість поживного середовища, на якому вони вирощуються.
Список використаних джерел
- Дідух Я.П. Популяційна екологія / Я.П. Дідух. – Київ: Фітосоціоцентр, 1998. – 192 с.
- Екологія: навчальний посібник / Н.М. Чернова, О.М. Билова. – Київ : Вища школа, 1986. – 230с.
- Маркіна Т.Ю. Особливості розведення рідкісних та зникаючих видів комах у лабораторних умовах / Т.Ю. Маркіна // Біологія та валеологія, 2014. – Вип. 16. – С. 37–46.
- Основи екології : навч. посіб. / О. М. Микитюк, В. В. Грицайчук, О. З. Злотін, Т. Ю. Маркіна. – 2-е вид., стер. – Х. : ОВС, 2004. – 144 c.
- Ткачев О.А. Бабочки в домашнем инсектарии / Ткачев О.А., Ткачева Е.Ю. – М.: "Аквариум ЛТД", 2001. – 64 с.
- Чернышев В.Б. Экология насекомых / В.Б. Чернышев. – 1996 – 304 с.
Нові проєкти і роботи




Навчальні програми



Банер сайту

Код банера:
